研 究
研究グループ/創造生理学グループ(三輪グループ)
プロジェクトテーマ
- 上皮Na輸送
- プロジェクトリーダー
青葉香代
- プロジェクトメンバー
青葉香代
- サブテーマ
1)表皮の細胞分化とENaCの発現の相互関係
2)発生過程における成体型表皮細胞の形成とENaCの発現の相互関係
3)上皮Na輸送に対するaldosterone, AVTの作用の解析
- 概要
上皮のNa再吸収(Na輸送)機構の発現メカニズムを研究している。実験材料には哺乳類の腎尿細管におけるNa再吸収のメカニズムの研究のモデル上皮である両生類の皮膚を用いている。両生類皮膚のNa輸送機構は幼生で未分化であり、変態過程で分化する。そこで幼生皮膚は本研究の目的に好都合と考えられた。
いままでに,
1) Na輸送の発現に甲状腺ホルモンは無関係である
2) アルドステロンで発現する
3) プロラクチンと成長ホルモンはアルドステロン作用を阻害する
ことを見いだした (AJP 268: C218, 1995; AJP 271: C1059, 1996; J Exp Biol 206: 1137, 2003)。
現在、Na輸送に関わる分子の一つ、上皮Na チャネル(ENaC)の発現を、ENaCのmRNA、ENaC蛋白、ENaC蛋白の活性化、のそれぞれのレベルで、表皮の細胞分化と関連づけて研究中である (J Comp Physiol 176; 287, 2006)。
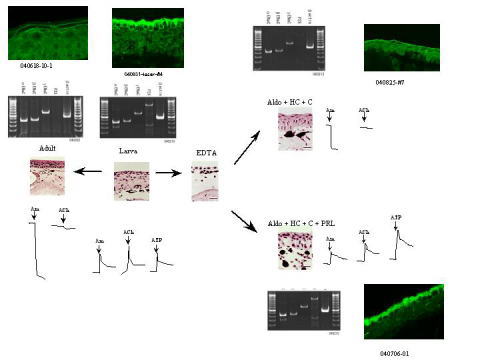
1)表皮の細胞分化とENaCの発現の相互関係
両生類の成体表皮は基底膜側から、胚芽細胞、有棘細胞、顆粒細胞、角化細胞で構成されている(ちなみに哺乳類でもこの構成は同じである)。これらの細胞は、胚芽細胞が分裂し、有棘細胞、顆粒細胞、角化細胞にと細胞分化することで形成されてゆく。最終的に角化細胞は剥がれ落ちる。表皮ではこの過程が繰り返される。成体両生類の表皮はENaCの関わるNa輸送を行っており、そのENaCは顆粒細胞の頂側に局在して機能していると考えられている。ENaCが関わる輸送であるかどうかは、輸送がENaCのinhibitorのamilorideという薬品で抑制されるか否かで判定する。
ENaCはこれら細胞分化の過程でどの細胞に発現し、いつ細胞膜(の頂側)に局在するのか。
例えば、有棘細胞が形成されると発現し、しかしまだ細胞内に留まっていて上皮輸送に関わることは無い。有棘細胞が顆粒細胞に分化する時に頂側への局在が起こる、というようなことが有るのか。あるいは、顆粒細胞になって初めてENaCの合成と頂側への局在が起こり、そして機能もするようになるのか。
2)発生過程における成体型表皮細胞の形成とENaCの発現の相互関係
両生類の表皮は、幼生と成体とで全く異なる細胞構成をしている。幼生では基底膜側から、基底細胞、skein細胞、apical細胞と呼ばれる細胞からなっている。幼生両生類の表皮はENaCの関わるNa輸送を行っていない(amilorideでinhibitされることが無い)。成体型の表皮細胞は広い意味での変態期に、基底細胞から成体型の表皮細胞を分化しうるような細胞(成体の胚芽細胞)ができ、そこから前に述べた成体型の表皮の細胞分化が起こるようになる(よってENaCの関わるNa輸送も成体型表皮が形成されてから起こる)。
無尾両生類の変態というと、通常は前肢が出現する時をもって変態の始まりであるという言い方をする(それで通じる)。学問的にはこの時期は、変態のclimax(最盛期)の始まりであり、変態そのものはもっと前から徐々に起こっている。しかし前肢の出現というのは非常に見た目に分かりやすい現象なので、また変態によって両生類は陸に上がることができるようになる(前肢が出ないことには陸に上がりようがない)という了解があるので、通常は前肢が出現する時をもって変態の始まりであると言う言い方をする。ウシガエルでは前肢が出現する時期を発生過程のTK stage表では, XXと呼んでいる。TKはstage表を作ったヒトの名の頭文字である。
ところで、前肢が出現する数日前の幼生の表皮は全体的には幼生型なのだがその中にすでに成体型の表皮細胞がすでに出現している。たとえば顆粒細胞はSt XVIIIですでに若干出現している(St XVIIでは出現していない)。この顆粒細胞の由来ははっきりしないが、この時期、基底細胞の一部から胚芽細胞が形成され、その一部の胚芽細胞から有刺細胞、顆粒細胞への分化のstartがすでに一部で始まってしまっているのかもしれない。いずれにせよ、St XVIIIからSt XXまでは6日ぐらいかかる。(St XI-XIXをprometamorphosis, St XX以降をClimaxと呼ぶことが多いがウシガエルのstage表をこしらえたTaylor and KollrosはStage XVIII以降をmetamorphic stageと一括しているのは先見の明かもしれない)。
発生過程で顆粒細胞ができると(すなわちSt XVIIIの顆粒細胞に)すでにENaCは発現しているのか。
裏返せば、ENaCの発現と顆粒細胞の分化は一致するのか(ENaCが発現していない顆粒細胞などというのがあるのか)。また、これらを調節する要因は何か?
以上が当面の課題である。
3)上皮Na輸送に対するaldosterone, AVTの作用の解析
AldosteroneはENaCのmRNA, ENaC 蛋白の発現を促進すると言われている。表皮は多層である。Aldosteroneは表皮を構成するどの細胞層のENaC mRNA, ENaC proteinの発現を促進するのか。
上皮Na輸送は vasopressin familyのhormoneでも調節を受ける。Vasopressin familyのhormoneの受容体には、V1-typeとV2-typeの2種がある。異なる受容体を介するarginine、 vasotocin (両生類におけるvasopressin familyのhormone)の作用の相違について研究している。
- 業績
【原著】
Kayo Fujimaki-Aoba, Kayoko Tanaka, Reiko Inomata, Philip JENSIK and Makoto TAKADA, (2014) α-ENaC in bullfrog embryo: expression in cement gland, gills, and skin. Cell and Tissue Reserch: 355(1),103-109.
Kayo Fujimaki-Aoba, Shinji Komazaki, Philip JENSIK, Shigeru Hokari and Makoto TAKADA, (2013) Larval bullfrog skin lacks amiloride-blockable epithelial transport because α-ENaC are within intracellular vesicles, not in apical plasma membrane, in epidermal apical cells. Acta Histochemica: 115,357-362.
Yuko Kaneko, Kayo Fujimaki-Aoba, Shu-Ichi Watanabe, Shigeru Hokari and Makoto TAKADA (2012) Localization of ENaC subunit mRNAs in adult bullfrog skin. Acta Histochemica: 114,172-176.
Makoto TAKADA, Kayo Fujimaki-Aoba and Shigeru Hokari (2011) Vasotocin- and mesotocin-induced increase in short-circuit current (SCC) across tree frog skin. J. Comp. Physiol. B: 181,239-248.
Makoto TAKADA, Kayo Fujimaki-Aoba and Shigeru Hokari (2010) Effects of arginine vasotocin and mesotocin on the activation and development of amiloride -blockable short-circuit current across larval, adult, and cultured larval bullfrog skins. J. Comp. Physiol. B: 180(3), 393-402.
Makoto TAKADA, Kayo Fujimaki and Shigeru Hokari (2008) Vasotocin has the potential to inhibit basolateral Na+/K+-pump current across isolated skin of tree frog in vitro, via its V2-type receptor/cAMP pathway. J. Comp. Physiol. B: 178, 957-962.
【学会発表】
青葉-藤牧香代・高田真理“ウシガエル皮膚における成体形質の発現時に変化する物質の同定” 第71回日本動物学会関東支部大会,東京,2019
菅理江・青葉-藤牧香代・高田真理“吸着行動をもちいた、ウシガエル幼生吸着器に発現するENaCの活性の検討” 第95回日本生理学会大会,高松,2018
菅理江・青葉-藤牧香代・高田真理“アフリカツメガエル幼生吸着器でのENaCの発現と 吸着行動における機能” 第94回日本生理学会大会,浜松,2017
Suge, R. and Takada, M “Function of amiloraide-blockable epithelial Na+ channel expressed in cement glands of young Xenopus laevis and bullfrog tadpoles” The 10th FENS Forum of Neuroscience, Copenhagen, Denmark, 2016, Jul.
菅理江・高田真理 “Function of amiloride-blockable epithelial Na+ channel on hanging behavior in young Xenopus laevis tadpoles” 日本動物心理学会第75回大会,東京,2015
Takada, M., Fujimaki-Aoba, K., Jensik, Philip J., Tanaka K., Shimomura, T., Inomata, R., Komazaki, S., Hokari, S., Cox, T. “Ontogeny of localization of epithelial sodium channel (ENaC) in bullfrog ectoderm”The 120th Annual Meeting of the Japanese Association of Anatomists & The 92nd Annual Meeting of the Physiological Society of Japan, Kobe, 2015
コンタクト埼玉医科大学生理学
場所:
基礎医学棟5F
520号室:三輪教授室
516号室:受付
連絡先:(受付)
TEL: 049-276-1152(内線2083)
FAX: 049-294-9961
〒350-0495
埼玉県入間郡毛呂山町毛呂本郷38